

Publicaciones SERIDA
Compuestos de naturaleza fenólica y actividad antioxidante de los brotes del pino (Pinus pinaster Aiton)
CAROLINA DE LA TORRE. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. delaorre85@gmail.com
JOSÉ FRANCISCO FUENTE. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. francisco@serida.org
MÓNICA MEIJÓN. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. mmeijon@serida.org
ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. ifeito@serida.org
SILVIA BAIZÁN. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. sbaizan@cetemas.es
ANGELO KIDELMAN DANTAS. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. kidelman@serida.org
JUAN MAJADA. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. jmajada@cetemas.es
VICTOR GRANDA. Universidad de Oviedo. Departamento Biología de Organismos y Sistemas. victorgrandagarcia@gmail.com
ANA RODRÍGUEZ. Universidad de Oviedo. Departamento Biología de Organismos y Sistemas. aralonso@uniovi.es

Figura 1.- Crecimiento vegetativo de Pinus pinaster, formación de un nuevo verticilo.
Los antioxidantes
Las barreras de defensa que establecen las plantas para protegerse de los agentes nocivos son diferentes a las que nosotros, u otros animales, somos capaces de formar. A los animales, incluido el hombre, la evolución les dotó de un sistema de defensa muy eficaz, con células o tejidos específicos a tal efecto. En las plantas cada célula actúa casi como un organismo autónomo que desarrolla sistemas de protección completos, aunque esto no quiere decir que no tengan tejidos con función protectora, como pueden ser las cortezas de los árboles, donde la concentración de sustancias de defensa suele ser más alta, actuando además como barrera física. Para optimizar y por tanto simplificar los mecanismos de defensa de forma que puedan ser repetidos a nivel de cada célula, las plantas invierten un gran esfuerzo en atenuar los efectos finales que causan la mayor parte de las agresiones ambientales a las que se ven sometidas, determinantes de lo que se engloba con el término estrés oxidativo.
El estrés oxidativo describe un estado de daño causado por la producción de especies reactivas de oxígeno (ROS) en una cantidad tal que excede a su destrucción o retirada; este exceso de ROS puede reaccionar con proteínas, ADN y otras moléculas esenciales para los organismos. El daño oxidativo también afecta al ser humano, relacionándose directamente con el envejecimiento y con algunos tipos de cáncer.
La ingesta de agentes antioxidantes provenientes del reino vegetal es una práctica muy recomendable y una forma de aprovechar para nuestro propio beneficio este recurso natural al cual podemos acceder a través de dietas ricas en frutas y verduras, como la mediterránea; la ingesta moderada de vino, por su contenido en resveratrol; los suplementos alimenticios (generalmente derivados de frutas del bosque) e incluso a través de la aplicación de cremas de origen vegetal, capaces de eliminar radicales libres.
Pero los vegetales de los que podemos extraer este recurso tan valioso no se reducen a aquellos de uso habitual en nuestra alimentación, sino que también es posible obtenerlos de las especies forestales. Éstas sintetizan una enorme variedad de compuestos, muchos de ellos aún desconocidos, que constituirán sin duda un nuevo modelo de bioeconomía en un futuro próximo. Este modelo se basará en el uso de los recursos biológicos renovables, mediante el desarrollo de bioprocesos y fabricación eco-eficiente de productos.
Actualmente, además de los productos forestales convencionales, ya existen más de 500 bioproductos procedentes de los árboles en el mercado. No obstante, la relevancia en la economía de estos bioproductos es difícil de cuantificar debido a que las estadísticas realizadas a día de hoy hacen referencia al segmento de mercado y no al origen del producto. Una parte importante de estos productos proceden de coníferas (pinos, abetos…), los cuales se caracterizan por tener un alto poder antioxidante y han sido evaluados para usos muy diversos, como alimentación, cosmética o usos clínicos. Podemos citar a modo de ejemplo el Pycnogenol® y el Enzogenol®, ambos producidos a partir de dos especies de pinos muy conocidas en nuestra región: Pinus pinaster (o pino del país) y P. radiata (o pino insigne), respectivamente .

Fifura 2.- Crecimiento de primavera de Pinus pinaster
Dado el creciente interés en los bioproductos y la importancia del metabolismo secundario, el estudio del contenido de estos compuestos de naturaleza fenólica presenta un doble interés: conocer los compuestos que podrían ser susceptibles de ser empleados como bioproductos y, por otra parte, utilizar esta información enprogramas de mejora genética de la especie, ya que nos permitirá tener caracterizados y conservar los materiales adaptados a condiciones ambientales diferentes.
El pino del país
En la “Estrategia Española para la Conservación y el Uso Sostenible de los Recursos Genéticos Forestales (ERGF)”, aprobada en primera instancia por el Comité Nacional de Mejora y Conservación de Recursos Genéticos Forestales (11 de Mayo de 2006, La Laguna, Tenerife) ya se indica la importancia que tienen para nuestros recursos forestales los grandes cambios que se están produciendo a nivel ambiental, los cuales podrán ocasionar respuestas a nivel biótico, siendo las nuevas condiciones ambientales inciertas y la respuesta biológica igualmente imprevisible.
La conservación de los recursos genéticos, por tanto, debe ir dirigida a mantener y preservar, en la medida de lo posible, aquellos procesos que faciliten la evolución bajo las nuevas condiciones ambientales de los ecosistemas, mediante el mantenimiento de los factores que intervienen en la estructuración de la diversidad genética de las especie.
En la ERGF se indica que la conservación y mejora se puede aplicar a distintos niveles, por lo que se puede distinguir entre prioridades a nivel de especie, de poblaciones y de genotipos. Para la mayoría de especies forestales, algunas poblaciones tienen un grado de amenaza suficiente para justificar la puesta en marcha de programas de conservación. El punto de partida para la mejora suelen ser los materiales vegetales de algunas regiones de procedencia que han demostrado su superioridad para los caracteres de interés. El Anexo I establece las especies que en el momento actual se consideran prioritarias como objeto de la ERGF. En la estrategia se han incluido las especies que son utilizadas en actividades de reforestación o restauración (incluidas en el RD283/2003 o en decretos autonómicos), las incluidas en el programa de conservación europeo EUFORGEN, aquellas con programas de mejora o con interés etnoagrario, y las que son objeto de gestión forestal. En la ERGF se justifica además la necesidad y se articulan los mecanismos para el establecimiento de Planes Nacionales de Mejora Genética Forestal. La gestión forestal se enfrenta a nuevos escenarios (cambio global, nuevas demandas sociales, plagas, etc.), para lo que precisa de materiales forestales de reproducción adecuados a estos fines.
Debido a este interés, el Principado de Asturias en colaboración con el CIFOR INIA, ha seleccionado P. pinaster (Figuras 1, 2 y 5), nuestra conífera más importante, como especie “modelo” para poner en marcha un programa de conservación y mejora de ámbito nacional incluyendo poblaciones naturales y materiales seleccionados de casi treinta procedencias distintas. Dentro de este proyecto, y a lo largo de los últimos 8 años, el Programa de Investigación Forestal (PIF), constituido por personal del Servicio Regional de Investigación y Desarrollo Agroalimentario (SERIDA) y del Centro Tecnológico Forestal y de la Madera de Asturias (CETEMAS), ha establecido una colección clonal, que es un referente internacional de materiales genéticos estructurados en una población base de mejora y/o materiales que cubren toda la diversidad genética poblacional en P. pinaster (Figura 3).
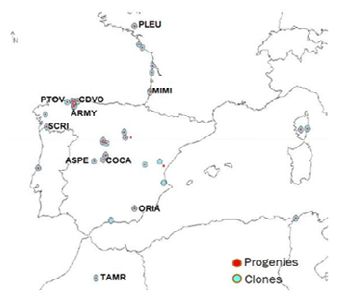

Figura 3. - Parcela experimental de La Mata (Grado) y origen de las procedencias de Pinus pinaster empleadas en este estudio. PLEU (Pleucadec), MIMI (Mimizan), PTOV (Puerto de Vega), CDVO (Cadavedo), ARMY (Armayán), SCRI (San Cipriano), COCA (Coca), ASPE (Arenas de San Pedro), ORIA (Oria), TAMR (Tamrabta).
Comparar materiales de origen tan diverso, abarcando desde la zona central de Francia hasta Marruecos, en una misma ubicación geográfica (parcela experimental de La Mata, Grado, 43° 32´N 7°00´W, 65 m) permite observar y analizar la gran diversidad de respuesta de los pinos bajo las mismas condiciones ambientales. A modo de ejemplo de la diversidad de respuestas, en la simple observación de visu de la parcela, podemos ver que los brotes anuales que forman los pinos en el crecimiento de primavera, tras la parada invernal, presentan una coloración diferente y aparentemente relacionada con el origen de la procedencia.
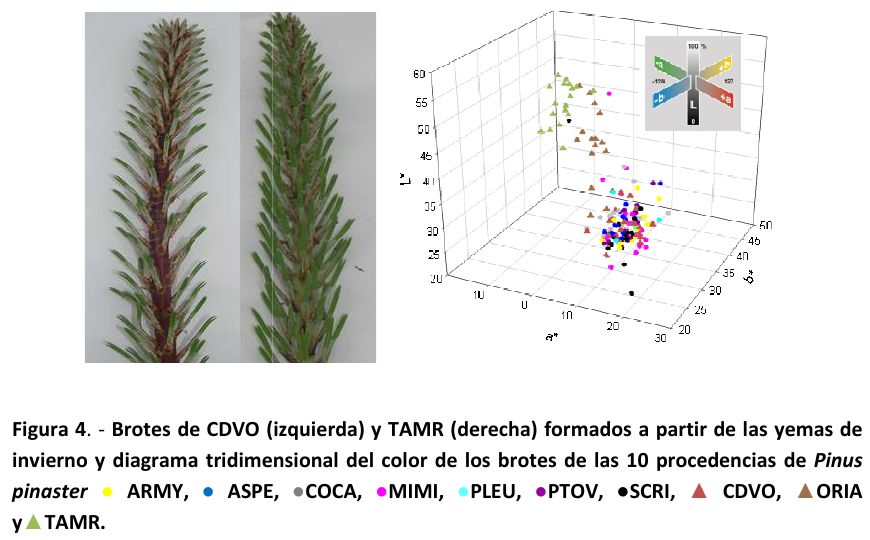
Como podemos ver en la Figura 4, donde se representan los parámetros del espacio de color CIE L*a*b* (CIELAB, modelo cromático usado para describir todos los colores que puede percibir el ojo humano), los triángulos verdes, que corresponden a la procedencia más al sur (TAMR, Marruecos), son los que se sitúan en la parte alta del gráfico de color seguidos, inmediatamente por debajo, de los triángulos marrones que representan la procedencia más al sur de la Península Ibérica (ORIA, Almería). Esta posición en el gráfico indica que estos materiales tienen un valor de la componente L* alto, o lo que es lo mismo, que son más claros; la componente a*, más negativa, indica que prima el verde sobre el rojo y la b*, más elevada, muestra una tendencia al amarillo.
Pigmentación
El color en las plantas depende de las concentraciones en que se encuentran los pigmentos y de las transformaciones a las que éstos se ven sometidos. Los pigmentos más importantes en el reino vegetal son las clorofilas, que dan el color verde característico de la mayor parte de los órganos aéreos de las plantas, tales como las hojas; los carotenoides, que dan los tonos amarillos o anaranjados, como el color de las zanahorias; las antocianinas, que confieren los tonos rosas, púrpuras, azules y rojos de muchas flores y frutos, y los flavonoides, pigmentos amarillos que dan color a muchos vegetales.
A pesar de que a cada pigmento le asignemos un color, la coloración final no siempre depende de la presencia de uno de ellos o de su concentración, pues además de que nunca están de forma aislada, el medio en que se encuentran condiciona el color final. Un claro ejemplo, a este respecto, lo constituyen los trabajos de mejora genética encaminados a obtener rosas azules. Se han conseguido transformar rosas con el gen del enzima responsable de la síntesis de la antocianina, delfinidina, que da el color azul a las petunias. Sin embargo, aún no se ha logrado el objetivo concreto, la “creación” del ejemplar azul, por la imposibilidad de modificar la acidez (pH) del contenido celular de los pétalos para que este pigmento sea realmente azul.
Un dato importante es que estas moléculas, además de su función como pigmentos propiamente dichos, están íntimamente relacionadas con la fisiología de la planta. Así, las clorofilas permiten realizar la fotosíntesis. Para otros pigmentos la funcionalidad no es tan clara, de ahí que muchos de ellos se agrupen dentro de los llamados metabolitos secundarios.
Hasta hace algún tiempo los metabolitos secundarios se solían considerar sustancias de desecho para el vegetal, carentes de una función fisiológica definida. En la actualidad se sabe que, si bien no tienen una trascendencia tan grande como los metabolitos primarios, sí son de gran importancia para el organismo en su relación con el entorno y supervivencia. Dentro de estas relaciones, la función más evidente quizá sea su papel como atrayentes de insectos, como ocurre en la polinización, o en su relación con otros vegetales, como ocurre con las sustancias alelopáticas, responsables de que una planta inhiba el desarrollo de otra.
Pero además, los metabolitos secundarios actúan como protectores frente a las agresiones producidas por infecciones, caso de las fitoalexinas, o a los depredadores, caso de los disuasorios nutritivos o alimentarios. Dentro de esta actividad protectora, los pigmentos también confieren fotoprotección y así, al igual que la melanina nos protege de los rayos del sol, en los vegetales, las antocianinas por ejemplo, les protegen de la radiación y fundamentalmente de la fracción ultravioleta. Bajo este punto de vista, cabría esperar que las procedencias del sur tuviesen una mayor coloración que las del norte, sin embargo nos hemos encontrado que en los pinos no es así, y que tampoco lo es en otras especies vegetales.

Figura 5.- Crecimiento de verano de Pinus pinaster
La acumulación de antocianinas, incluso en flores y frutos, tiene lugar de forma más acusada en las procedencias de latitudes más al norte o de zonas de mayor altitud. Este hecho es lo que nosotros pudimos constatar en pino (Tabla 1) y, según explican algunos autores refiriéndose al contenido en frutos, parece deberse al hecho de que en estas latitudes o altitudes la radiación del sol y las horas de sol son elevadas en momentos en los que la temperatura aún es baja.
Las plantas utilizan la energía del sol para sintetizar sus propios nutrientes, de forma que la protección frente al mismo debe de orquestarse muy eficientemente, compaginándola con su uso como recurso, y por tanto es un proceso mucho más complejo del que ocurre en los animales. Los resultados que hemos obtenido, al igual que los obtenidos por otros autores, parecen indicar que cuando la temperatura y los recursos hídricos o nutricionales son suficientes como para mantener una actividad fotosintética elevada no resulta nociva la radiación solar y, por tanto, las procedencias del sur tendrán menor contenido en antocianinas. Sin embargo, cuando la temperatura no es suficiente y la eficiencia fotosintética es baja, el exceso de luz solar genera la formación de los radicales libres que provocarán el daño oxidativo y, por ello, las procedencias de las latitudes norte muestran esa predisposición a acumular un mayor contenido en antocianinas.
El interés de estos estudios a nivel práctico es evidente, pues nos indicarían que las frutas de las latitudes norte nos aportarían una mayor concentración de antioxidantes que las del sur para una misma especie.
Tabla 1. - Valores medios (± error estándar) y coeficiente de variación (CV) del contenido en fenoles totales (mg EAG g-1 PS), flavonoides (mg EQ g-1 PS), clorofila a, clorofila b, carotenoides y antocianinas (µmoles g-1 PS) para las procedencias de Pinus pinaster CDVO, ORIA y TAMR. Letras distintas muestran diferencias significativas entre procedencias. Test de Tukey P< 0,05.
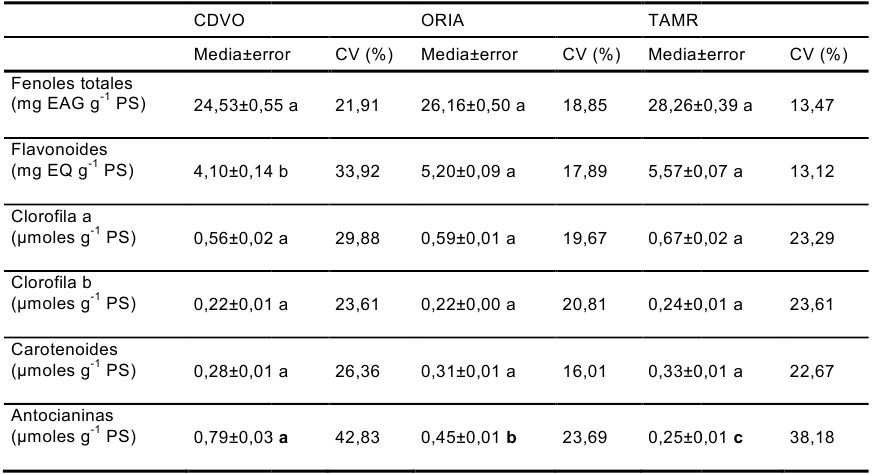
Además de las antocianinas hemos analizado otros compuestos con capacidad antioxidante, algunos de ellos coincidentes con los que forman parte del Pycnogenol®, no apreciándose diferencias, según la procedencia, a excepción del contenido en flavonoides totales, que sería favorable a las procedencias del sur. Las procedencias del sur, por tanto, a pesar de su menor coloración, no estarían tampoco exentas de una protección frente al daño oxidativo, que como ya hemos comentado, puede ser desencadenado por cualquier factor estresante, como ocurre por ejemplo con la falta de agua. Basándonos en estos resultados, podemos decir que tanto unas procedencias como otras son capaces de acumular algún tipo de antioxidante y que lo único que parece cambiar es la naturaleza de los componentes que les aportan esa protección, como resultado quizá de una adaptación a unas condiciones ambientales muy diferentes. También podríamos concluir que todas ellas serían válidas para la obtención de antioxidantes, si bien, los datos obtenidos no son un reflejo directo del potencial de esta especie ya que se analizaron los brotes y no la corteza. No obstante, sí se podrían considerar como un buen indicador ya que estos brotes son en realidad el origen de los diferentes tejidos de los tallos, entre los que se encuentra la futura corteza.
La corteza de los árboles es el tejido que se utiliza para la obtención de bioproductos por su papel de tejido de protección y por tanto acumulador de mayor contenido de antioxidantes. La elección de los brotes para este análisis se debe a su mayor sensibilidad al estrés oxidativo por ser tejidos juveniles y, en cuanto a la época del año, se seleccionó el crecimiento de primavera por ser el momento más determinante para la planta en cuanto a este tipo de estrés, ya que en esta época hay suficientes horas de insolación con temperaturas aún bajas.
El residuo obtenido de las extracciones muestra la diferente coloración característica de los pigmentos más abundantes (roja para las antocianinas, abundantes en la procedencia CDVO y amarilla para los flavonoides, más abundantes en ORIA y TAMR, Figura 4) y dio un rendimiento en pigmentos cercano al 40 % del peso seco extraído. Este rendimiento es muy aceptable si tenemos en cuenta que aunque la época del año sea favorable, el tejido extraído no es el más idóneo para la obtención de bioproductos, como hemos señalado previamente.

Figura 6. - Residuo obtenido tras la extracción de los pigmentos de los brotes de tres procedencias de Pinus pinaster, CDVO, ORIA y TAMR.
Como conclusión podemos decir que esta especie (Pinus pinaster) desarrolla un sistema de defensa frente al estrés oxidativo que presenta variabilidad dependiente de la procedencia. Que esta variabilidad parece relacionada con la latitud, al igual que ocurre en otras especies, y que da lugar a un carácter fenológico valorable en el espacio de color CIE L*a*b. Y, por último, se constata el potencial de esta especie como fuente de bioproductos, pues a pesar de no haber sido utilizado el tejido diana idóneo para los análisis bioquímicos, el rendimiento obtenido está dentro de lo esperable.
Agradecimientos
Este trabajo forma parte del Proyecto Fin de Máster que Silvia Baizán presentó en el mes de julio de 2013, dentro del Máster en Biotecnología Aplicada a la Conservación y Gestión Sostenible de Recursos Vegetales y de la Tesis Doctoral que Carolina de la Torre se encuentra elaborando en la actualidad dentro del Proyecto RTA 2010 00120, Subprograma de Proyectos de Investigación Fundamental Orientada a los Recursos y Tecnologías Agrarias.
Referencias bibliográficas
Alía R., Martín S. 2009. Guía técnica para la conservación genética y utilización del pino negral (Pinus pinaster) en España. Foresta. Madrid. España. 2 pp.
Baizán S. 2013. Determinación de la composición fenólica y actividad antioxidante en brotes de clones de Pinus pinaster procedentes de tres orígenes contrastantes. Máster en Biotecnología Aplicada a la Conservación y Gestión Sostenible de Recursos Vegetales. Univ. Oviedo.
Brzeziñska E., Koz³owska M. 2008. Effect of sunlight on phenolic compounds accumulation in coniferous plants. Dendrobiology 59: 3-7
Chalker-Scott L. 1999. Environmental significance of anthocyanins in plant stress responses. Photochemistry and Photobiology 70(1): 1-9.
Gould K. S., Quinn B. D. 1999. Do anthocyanins protect leaves of New Zealand native species from UV-B? New Zeal J Bot 37: 175-178.
Jaakola l., Hohtola A. 2010. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ 33: 1239–1247.
Lätti A. K., Riihinen K. R., Kainulainen P. 2008. Analysis of anthocyanin variation in wild populations of bilberry (Vaccinium myrtillus L.) in Finland. J Agr Food Chem 56: 190-196.
Nozzolillo C., Isabelle P., Andersen Ø. M. Abou-Zaid M. 2002. Anthocyanins of jack pine (Pinus banksiana) seedlings. Can J Botany 80: 796–801.
Oleszek W., Stochmal A., Karolewski P., Simonet A. M. , Macias F. A., Tava A. 2002. Flavonoids from Pinus sylvestris needles and their variation in trees of different origin grown for nearly a century at the same area. Biochem Syst Ecol 30: 1011–1022.
Treutter D. 2006. Significance of flavonoids in plant resistance: a review. Environ Chem Lett 4: 147–157.
| Ficha Bibliográfica | |
|---|---|
| Título | Compuestos de naturaleza fenólica y actividad antioxidante de los brotes del pino (Pinus pinaster Aiton) |
| Autor/es | CAROLINA DE LA TORRE. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. delaorre85@gmail.com JOSÉ FRANCISCO FUENTE. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. francisco@serida.org MÓNICA MEIJÓN. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. mmeijon@serida.org ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación Forestal. ifeito@serida.org SILVIA BAIZÁN. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. sbaizan@cetemas.es ANGELO KIDELMAN DANTAS. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. kidelman@serida.org JUAN MAJADA. Centro Tecnológico y Forestal de la Madera de Asturias. Programa de Investigación Forestal. jmajada@cetemas.es VICTOR GRANDA. Universidad de Oviedo. Departamento Biología de Organismos y Sistemas. victorgrandagarcia@gmail.com ANA RODRÍGUEZ. Universidad de Oviedo. Departamento Biología de Organismos y Sistemas. aralonso@uniovi.es |
| Año Publicación | 2014 |
| Área | |
| Revista/Serie | Tecnología Agroalimentaria. Boletín informativo del SERIDA |
| Referencia | N. 13. pp. 27-33 |
| Formato | |
| Depósito Legal | 2.617/1995 |
| ISSN | 1135-6030 |
| ISBN | |
| Ver/Descargar | |