

Publicaciones SERIDA
Minipropagación de plantas forestales
ÂNGELO KIDELMAN DANTAS. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. kidelman3@hotmail.com
JUAN MAJADA. Centro Tecnológico y Forestal de la Madera (CETEMAS). jmajada@cetemas.es
MARTA CIORDIA. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. mciordia@serida.org
ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. ifeito@serida.org
Introducción
La reproducción asexual vegetal es posible debido a que cada célula de la planta contiene la información genética necesaria para generar la planta entera. Esta propiedad para generar un organismo completo se denomina totipotencia (Hartmann y Kester, 1998).
Ante la ausencia de sistemas económicamente viables por métodos biotecnológicos en ciertas especies, la optimización a nivel comercial del enraizamiento de esquejes en especies como el eucalipto se ha conseguido, especialmente, con el desarrollo de la técnica de miniesquejado (Xavier y Wendling, 1998; Wendling et al., 2000), definida como la propagación vegetativa a través de minisetos, no rejuvenecidos previamente “in vitro”, pero gestionados intensivamente mediante podas (Wendling y Xavier, 2001). Esta técnica posibilita considerables ganancias, principalmente en el aumento de los índices de enraizamiento, reducción del tiempo de formación de las plantas por el uso de propágulos con mayor grado de juvenilidad y facilita una mejor funcionalidad del sistema radical (Titon et al., 2002).
La técnica de miniesquejado cuenta con un aliado imprescindible que es el uso de los reguladores del crecimiento vegetal, principalmente auxinas, que incrementan el porcentaje de esquejes que forman raíces, el número y calidad de las raíces producidas por esqueje, acelera la iniciación de éstas (Hartmann y Kester, 1998) y uniformiza el enraizamiento (MacDonald, 1995; Hartmann y Kester, 1998). Según Azcon-Bieto y Talon (1993), la mayor probabilidad de formar raíces adventicias en los esquejes sometidos a diferentes concentraciones de AIB estaría relacionada con el efecto estimulador de la diferenciación en las raíces provocado por las auxinas.
Así, el miniesquejado, como técnica de propagación vegetativa y asociada al uso de reguladores de crecimiento, ha demostrado ser un método eficiente para la propagación de muchas especies forestales, existiendo protocolos de producción comercial para especies como el eucalipto (Eucalyptus spp.) (Campinhos, 1987), el pino insigne o de Monterrey (Pinus radiata D.), la pícea común o abeto rojo (Picea abies (L.) H. Karst) y el pino taeda o pino toro o del Norte de Carolina (Pinus taeda L.) (Ritchie, 1991), entre otras. El Programa de Investigación Forestal ha contribuido con la puesta a punto de protocolos para varias especies, de las cuales, las dos más relevantes ya han sido objeto de publicación: Pinus pinaster Ait. (Majada et al., 2011; Martínez-Alonso et al., 2012) e híbridos artificiales de Castanea sativa Mill. (Dantas et al., 2016). El trabajo que ahora se presenta es un compendio de los resultados de enraizamiento más destacados en los últimos años en este Programa para el pino pinaster (P. pinaster Ait.), el cerezo (Prunus aviun L.), una selección de híbridos artificiales de C. sativa Mill. y el tejo (Taxus baccata L.).
La necesidad de recurrir a sistemas de multiplicación asexual surge a partir del momento en que contamos con materiales vegetales de calidad genética superior. En Asturias, se promueven el interés por estos materiales superiores a partir de 2003, cuando se inicia la andadura del Programa de Investigación Forestal, que plantea la necesidad de multiplicar especies forestales de interés por su alto valor añadido, tal como el pino y el castaño; o como base de una posible diversificación en la producción forestal, como es el caso del cerezo, o de conservación de especies emblemáticas, caso del tejo.
En la “Estrategia Española para la Conservación y el Uso Sostenible de los Recursos Genéticos Forestales”, se define que la conservación de los recursos genéticos debe estar dirigida a mantener y preservar aquellos procesos que faciliten la evolución bajo las nuevas condiciones ambientales de los ecosistemas, mediante el mantenimiento de los factores que intervienen en la estructuración de la diversidad genética de las especies. Los planes de actuación recogidos en el marco de la estrategia integran el desarrollo de herramientas y medidas para abordar aspectos concretos de la conservación y el uso sostenible de los recursos genéticos forestales.
El interés en minipropagar en Asturias los materiales forestales seleccionados
El pino pinaster se consideró especie modelo para los primeros ensayos de miniesquejado ya que es la principal conífera del sur de Europa, con gran importancia en el mediterráneo occidental, donde ocupa extensas superficies y forma pinares multifuncionales. La madera y la resina son los dos usos más importantes de la especie, si bien también tiene un uso ambiental, al adaptarse a un gran abanico de condiciones agroclimáticas. “Puede encontrarse en ambientes muy diferentes: desde el nivel del mar hasta 2.100 m en el Alto Atlas (Marruecos); desde áreas con más de 1.400 mm de precipitación anual y sin temporada seca a otros con 350 mm y más de 4 meses secos. Las condiciones de suelo son variables: crece principalmente en suelos ácidos, pero también en suelos básicos y hasta en suelos arenosos y pobres, donde pocas especies con valor comercial pueden crecer” (Alía y Martín, 2009).
En el proyecto TREESNIP (V programa Marco) se caracterizaron 22 poblaciones y familias de P. pinaster de toda el área de distribución de la especie. Partiendo de los resultados de este proyecto y, liderado por el programa forestal de investigación del SERIDA (RTA2006-00084-00-00), se generó una población base para un programa de mejora genética de ámbito estatal y se abordaron diversos aspectos en relación a la tolerancia al estrés hídrico de la especie. A partir de entonces, se establecen dos colecciones: Colección 1; con cinco genotipos clonados por familia y Colección 2; clones con un genotipo por familia, presentes en la población base de campo (Fig. 1. Colecciones de Referencia “CLONAPIN”).

Figura. 1. Vista general de plantas madre (izquierda), detalle de las mismas (centro) y planta clonada (derecha) de pino pinaster.
El cerezo es, y ha sido, muy apreciado como componente de la diversidad de los ecosistemas forestales de toda Europa, donde se extendió procedente de Asia occidental, acompañando al hombre desde tiempos remotos. A nivel mundial, se encuentra difundido por numerosas regiones y países con clima templado (Rusell, 2003), demostrando ser una especie con una gran capacidad de adaptación a distintas condiciones climatológicas, ya que lo podemos encontrar desde Gran Bretaña al Cáucaso y desde el norte de África a los países escandinavos (López-González, 2001; Montero et al., 2003). En España, esta especie se distribuye sobre todo por la mitad norte peninsular, siendo un árbol frecuente en Galicia, especialmente en la mitad oriental de la región (Álvarez et al., 2000), si bien Las Regiones de Identificación y Utilización de material forestal de reproducción (RIU’s) números 7 y 9, pre-Pirineo y bosques vasco-navarros, son las que presentan mayor presencia de cerezos (García del Barrio et al., 2001). Es una especie heliófila, que suele hallarse naturalmente en los bordes de masas, caminos, arroyos, etc., y cuyo interés, desde un punto de vista paisajístico, radica en su floración espectacular y en la coloración otoñal de sus hojas. También resalta su valor natural, por servir de alimento para la fauna silvestre y, desde un punto de vista forestal, por la posibilidad de producir madera de excelente calidad.
En el marco del proyecto INIA SC98-061-C3-2, 1998-2001 se realizaron actividades de prospección y selección de árboles superiores en Galicia, Asturias, Navarra, País Vasco y Castilla y León, basada en criterios de selección fenotípica de aptitud forestal y se establecieron varios huertos semilleros, mediante injertado, uno de ellos en Asturias (Ciordia et al., 2015). Además, se clonaron “in vitro” algunos de los árboles élite seleccionados en campo. Posteriormente se pondrá a punto la multiplicación por miniestaquilla (Fig. 2).
.jpg)
Figura. 2. Cerezo clonado por miniestaquilla.
El castaño se encuentra ampliamente extendido en China, Japón y Europa. En Asia, destacan las especies Castanea mollissima Blume y Castanea crenata Sieb y Zuc. y en Europa, la especie C. sativa Mill. es la mayoritaria. En los últimos años se fomentó la expansión de híbridos interespecíficos con especies asiáticas tolerantes al chancro (Chryphonectria parasitica) y a la tinta (Phytophthora spp.) (Miller et al., 1996) y, actualmente, también a la avispilla (Dryocosmus kuriphilus). España cuenta con unas 440.000 ha de castaño, según el Mapa Forestal Nacional, repartido por casi todas las CCAA en mayor o menor grado, pero especialmente en Asturias, Lugo, Ourense, León y Salamanca. La superficie de cultivo como castaño de fruto es de unas 111.000 ha, con las mayores superficies en Ourense y Lugo, según datos de la red Estatal del Castaño. El desarrollo de la Norma Nacional de Clasificación Estructural para la madera de esta especie por parte del Centro Tecnológico Forestal y de la Madera de Asturias es importante para la selección de ejemplares por calidad de madera y para el desarrollo de una industria nacional. Así mismo, respecto al castaño de fruto, la iniciativa promovida como Grupo Operativo Nacional, desde las CCAA donde el castaño está más representado (Galicia, Asturias, Castilla-León, Cataluña, Andalucía y Extremadura), y en la que participa el SERIDA, será fundamental en la promoción de esta especie como cultivo.
Actualmente, el Catálogo Nacional de Materiales de Base dispone de materiales forestales de reproducción de castaño de semilla (identificada y seleccionada) y de aquellos del programa de mejora iniciado por Urquijo y Vieitez y conservados, caracterizados y reproducidos por el Centro de Investigaciones Forestales de Lourizán (Xunta de Galicia), así como la colección que TRAGSA ha desarrollado, que incluyen también resistencia a tinta y que han sido clonados mediante micropropagación (Cuenca et al., 2009).
En Asturias, considerando el castaño para uso forestal, se dispone de cinco fuentes semilleras, además de una plantación de material clonal de híbridos interespecíficos (Fig. 3) y de Una Población Base de progenies de C. sativa (Ciordia et al., 2015). Respecto al castaño de fruto, se mantiene una colección activa de germoplasma en las instalaciones del SERIDA, que incluye los principales cultivares de la región, registrándose recientemente 11 de ellos en la Oficina Española de Variedades Vegetales del Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente (BOE nº 60, de 11 de marzo de 2017).
.jpg)
Fig. 3. Material clonal de híbridos de castaño interespecíficos.
Establecidos los protocolos generales para la minipropagación vegetativa de materiales forestales de interés económico para la región, surgió la posibilidad de ampliar el marco de actuación a una especie cuyo objetivo, en el momento de plantear la actuación, era completamente diferente al de las especies ya mencionadas. Sin embargo, la conservación de especies dentro de un marco de protección del medioambiente o, como en este caso, de interés histórico-cultural, no es un aspecto menos relevante desde el punto de vista social, por sus particularidades como ejemplares únicos, caso de los tejos centenarios catalogados como tal. Con este planteamiento y, en colaboración con la Universidad de Oviedo-Fundación Valdés Salas y EDP, se hizo un trabajo para recuperar y clonar los tejos asturianos catalogados como Sitios Históricos, doce concretamente, con el objetivo de establecer un arboretum con todos los ejemplares.
El tejo es una especie de gran relevancia en todo el territorio asturiano, y si bien cuando se iniciaron los trabajos de multiplicación vegetativa su protección se incluía sólo bajo la figura de Árbol Monumental, recientemente se ha ampliado su rango de protección patrimonial a través de una nueva categoría denominada Sitio Histórico, que engloba la protección no sólo de los árboles sino de su entorno (BOPA nº 239, de 16 de octubre de 2017). La ley define un Sitio Histórico como un lugar vinculado a acontecimientos de interés histórico, a tradiciones populares o a creaciones culturales relevantes, permitiendo su protección a través de su declaración individualizada como Bien de Interés Cultural (BIC), lo que les otorgar el máximo nivel de protección que busca salvaguardar uno de los legados más significativos de la historia, cultura y tradición de Asturias. La mayoría de los tejos están representados en ambas figuras de protección, y todos ellos son árboles centenarios o milenarios, poderosos y longevos.
Material vegetal de ejemplares únicos de Sitios Históricos catalogados por sus particularidades como tal: la Ceñal en la Collada (Siero), Santa Coloma (Allande), Santibáñez de la Fuente (Aller), Rozadas (Boal), Montoubu (Belmonte de Miranda), Martul (Villanueva de Oscos), Perueño (Quirós), Arangas (Cabrales), San Martín del Mar (Villaviciosa), Cenero (Gijón), Abamia (Cangas de Onís) y San Martín de Salas (Salas) (Fig. 4), se recogió para su multiplicación y la creación de un banco clonal con el propósito de mantener el legado histórico.


Figura 4. Imagen del tejo de San Martín, Salas (izquierda) y detalle de ramillos terminales con frutos del año (derecha).
Optimización del miniesquejado de los materiales forestales de interés
Los programas de multiplicación vegetativa son una herramienta importante, tanto para los programas de mejora genética, como para incrementar la competitividad de las industrias de base forestal, con potencial para establecer plantaciones clonales con mayor productividad y calidad de madera (Borges et al., 2011), caso del eucalipto.
Pinus pinaster
La inducción del enraizamiento con tratamientos previos, como el frío, la definición de la edad ideal para enraizamiento de los esquejes, la optimización de la fertilización de las plantas madre para estimular la producción de esquejes, y su efecto sobre el enraizamiento, y el establecimiento del número máximo de ciclos de esquejado al año, fueron los objetivos planteados para el pino pinaster que permitieron desarrollar protocolos que, una vez puestos a punto, sirvieron y sirven para multiplicar el material seleccionado que se encuentra en los bancos de germoplasma del SERIDA (Fig. 5)

Figura. 5. Miniestaquillas de pino de varios clones de una misma procedencia
Durante los últimos años se consiguieron establecer protocolos de enraizamiento para el pino, con el almacenamiento de los esquejes en frío, a 4ºC, y a temperatura ambiente con estímulo externo de AIB, obteniéndose muy buenos resultados en ambos tratamientos, con un enraizamiento superior a los 92% y 97%, respectivamente (Tabla 1).
Tabla 1. Porcentaje de enraizamiento de diferentes especies forestales con su respectiva concentración de regulador de crecimiento vegetal (AIB) utilizando miniesquejes.
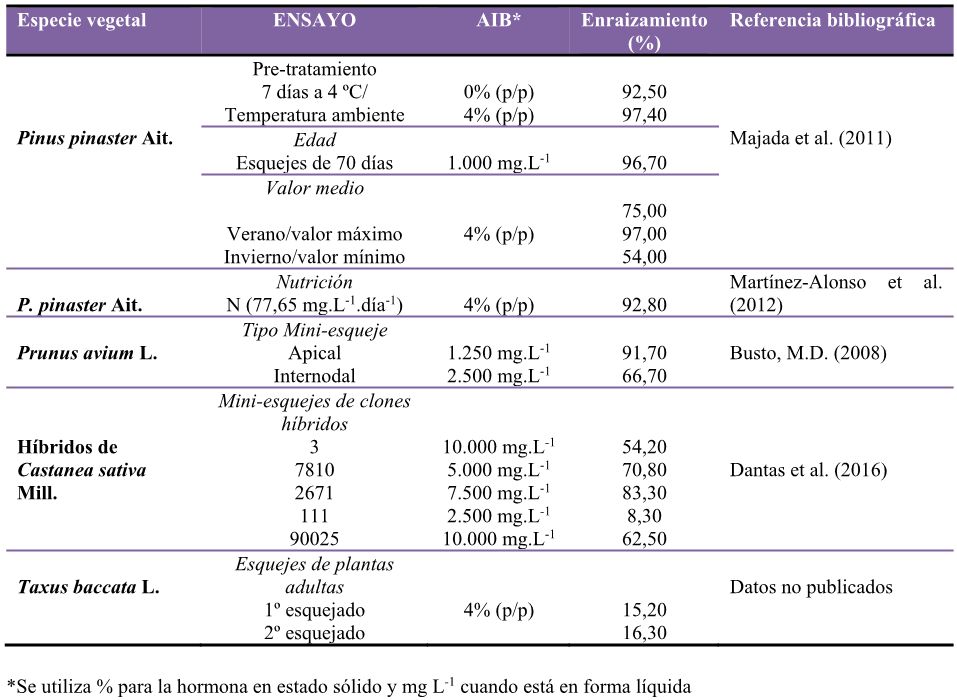
Por lo que respecta a la edad ideal para enraizar esquejes juveniles, nuestros resultados indican que el óptimo se encuentra en 70 días, asociado al aporte externo de reguladores de crecimiento (AIB), obteniéndose un porcentaje de enraizamiento superior al 96% (Tabla 1).
En cómputo anual, la media de enraizamiento de los esquejes se mantuvo en un 75%; no obstante, se observó una estacionalidad (Majada et al., 2011), con un descenso en el enraizamiento en el periodo invernal y alcanzando los valores más altos en el verano (Tabla 1).
La producción de esquejes es estimulada por un mayor aporte de nitrógeno aplicado a las plantas madre. Los datos obtenidos indican que se puede conseguir un incremento en la producción de esquejes suplementando la fertilización con un aporte de nitrógeno superior (Martínez-Alonso et al., 2012). Este ensayo se realizó con una selección de familias clonales de la Colección 1 de pino, anteriormente indicada. El efecto sobre el enraizamiento también fue objeto de estudio, y se consiguió un porcentaje superior al 92% (Tabla 1).
La tecnología de propagación clonal desarrollada ha facilitado, adicionalmente, generar material con el cual se ha establecido un dispositivo de ensayos clonales en ambientes contrastantes (Francia/España/Portugal) los cuales forman parte red de excelencia EVOLTRE; materiales que además han sido empleados en ocho proyectos de investigación de convocatorias de concurrencia competitiva, así como para el desarrollo de varias tesis doctorales, dos de ellas en nuestra comunidad autónoma (Velasco Conde T., 2011. Análisis de la respuesta a la sequía en Pinus pinaster Aiton a través de la evaluación de caracteres morfológicos, fisiológicos y genéticos. Universidad de Oviedo. Dirección: J. Majada e I. Aranda ; y Delatorre C., 2017. Bases fisiológicas y moleculares del crecimiento estacional de Pinus pinaster Aiton. Universidad de Oviedo; Dirección: A. Rodríguez e I. Feito), así como varios trabajos de investigación, además de los ya mencionados sobre propagación en esta especie (Delatorre et al., 2013; Gaspar et al., 2013; Cañas et al., 2015; Meijón et al., 2016).
Prunus avium
El cerezo fue otra de las plantas forestales en las que se consiguió establecer un protocolo de enraizamiento de esquejes juveniles con alta eficiencia, a la vez que se valoraron dos tipos de esquejes, apical e internodal (Fig 6), y diferentes concentraciones de AIB. Este trabajo se desarrolló a través de un Proyecto Fin de Carrera (Busto, M.D. 2008. Propagación clonal de material juvenil de Prunus avium L. Ingeniería Técnica Forestal, E.U. Universidad de Oviedo; Directores: J. Majada y A. Kidelman).
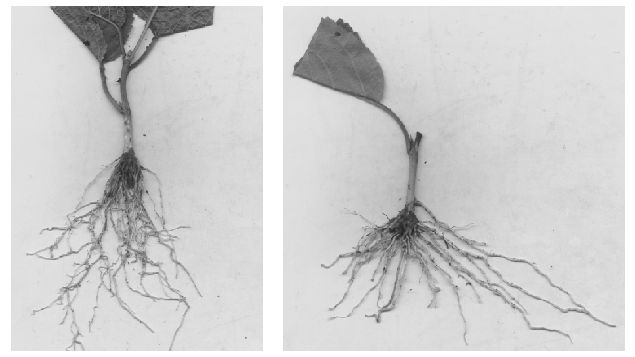
Figura. 6. Enraizamiento de segmento apical (izquierda) e intermodal (derecha) de Prunus avium.
Para el esquejado del cerezo se partió de materiales seleccionados por su vigor y calidad sanitaria, a partir de la fenología realizada en el huerto semillero ubicado en nuestra comunidad autónoma, dentro de los proyectos nacionales que dieron continuidad a esta línea (RTA2005-00057-C05-00 y RTA2011-00046-00-00). En los ensayos de optimización del enraizamiento se aplicaron diferentes concentraciones de solución nutritiva a la planta madre, variando la concentración de nitrógeno para estimular la producción de nuevos brotes y, una vez optimizada la fertilización, se obtuvieron esquejes juveniles con 3 a 4 yemas, diferenciando entre apicales e internodales, además de optimizar las dosis hormonales. Los resultados obtenidos muestran que los esquejes apicales necesitan una menor dosis hormonal (1.200 mg L-1 de AIB) y muestran una mayor capacidad de enraizamiento (91,7%) (Tabla 1). Este resultado, probablemente debido la juvenilidad de los esquejes, también podría ser la causa de la ausencia casi total de diferencias entre familias en cuanto a porcentaje de enraizamiento y supervivencia, para los esquejes apicales.
Castanea spp
Para el castaño, el protocolo se estableció para materiales híbridos dado su doble aptitud como material forestal y potencial uso como portainjertos. Se partió de planta madre adulta, multiplicada por acodo bajo, lo que hizo necesario un primer paso de rejuvenecimiento ya que el material adulto tiende a ser muy recalcitrante. La tasa de enraizamiento obtenida fue aceptable para la mayoría de los clones (de 54-83%), formando un buen sistema radicular (Fig. 7), pero fue preciso utilizar dosis elevadas de AIB (Dantas et al., 2016). La falta de uniformidad entre materiales y la baja eficiencia para el clon 111 indica que es necesario optimizar el proceso, siendo la fase de rejuvenecimiento la más crítica (Dantas et al., 2016).
La planta propagada se utilizó en la repoblación de dos obras de la Dirección de Política Forestal (Consejería de Desarrollo Rural y Recursos Naturales), Repoblación con castaño y abedul en el rodal A4. MUP 364 "Cordal de Peón" Villaviciosa, con cargo al fondo de mejoras y Actuaciones Forestales en el pinar de La Llama, Tineo (16.005-OBF).

Figura. 7. Esquejes de castaño enraizados
Taxus bacata
Se realizó la recogida de material vegetal, que consistió en brotes bajos con posición terminal y siempre con crecimiento del año, para favorecer que mantuvieran una cierta juvenilidad (Fig. 8). Para su enraizamiento se utilizó una única concentración de AIB al 4%, dosis alta teniendo en cuenta que se trataba de material adulto y una especie de difícil enraizamiento. El porcentaje de enraizamiento medio obtenido fue inferior al 17% (Tabla 1).

Fig. 8. Clones ya enraizados de los tejos seleccionados por su interés histórico.
Estos datos que, a priori pueden parecer muy bajos, no lo son tanto si se tiene en cuenta que es una especie recalcitrante para el enraizamiento; la plata es adulta, o más bien muy envejecida (ejemplares seculares), y en la que no se realizó ningún estudio sobre tratamientos previos para estimular el enraizamiento y posibilidad de rejuvenecimiento a través de cadenas de proliferación.
Considerando el conjunto de las especies revisadas en este trabajo reseñar, para finalizar, que todos los materiales propagados sirvieron, además de como material experimental para realizar trabajos científicos y formar parte de Bancos de Germoplasma activos en los que está directamente implicado el SERIDA, para potenciar la colaboración científico-técnica con otras entidades como Universidades, Centros de Investigación y Fundaciones de I+D+i en muy diversos campos de estudio como los relacionados con la genética, genómica, proteómica, metabolómica, fisiología, ecología, reforestación, selvicultura y gestión de estas especies.
Agradecimientos
Agradecemos especialmente la inestimable colaboración del personal de campo del Programa Forestal (SERIDA/CETEMAS), imprescindible para realizar todos estos trabajos. También agradecemos la colaboración del CIFOR-INIA y SERPA por el apoyo con los materiales vegetales de partida y la conservación de la planta madre. A los alumnos en prácticas que colaboraron en una labor tan tediosa y minuciosa como es el miniesquejado.
La financiación de estas actividades se llevó a cabo fundamentalmente con el apoyo de la Dirección General de Montes e Infraestructuras Agrarias de la Consejería de Desarrollo Rural y Recursos Naturales del Principado de Asturias.
Bibliografía
ALÍA, R. y S. MARTÍN. 2009. Guía técnica para la conservación genética y utilización del pino negral (Pinus pinaster). Foresta. Madrid. España.
ÁLVAREZ, P.; BARRIO, M.; DÍAZ, R. A.; HIGUERAS, J.; RIESGO, G.; REGUEIRO, A.; RODRÍGUEZ, R. J. Y VILLARINO, J. J. 2000. Manual de selvicultura de las principales especies de frondosas forestales para el norte de España. Prunus avium L. cerezo. Fecha de consulta 22/II/2008. Publicado en Internet, disponible en: http://www.agrobyte.com/agrobyte/publicaciones/frondosas/cap6_5.html
AZCON BIETO, J.; TALON, M. 1993. Fisiología y bioquímica vegetal. Interamericana. Madrid. España.
BOE. 2017. Orden APM/207/2017, de 7 de marzo. B.O.E. nº 60, de 11 de marzo de 2017, Sec. III, Anexo II, pág 18164.
BOPA. 2017. Decreto 61/2017, de 20 de septiembre. BOPA nº 239, de 16 de octubre de 2017.
BORGES S. R., XAVIER A., OLIVEIRA L. S., MELO L. A., ROSADO A. M. 2011. Enraizamento de miniestacas de clones híbridos de Eucalyptus globulus. Revista Árvore, 35 (3): 425-434.
CAÑAS R.A.; FEITO I.; FUENTE-MAQUEDA J.F.; AVILA C.; MAJADA J.; CANOVAS F.M. 2016. Transcriptome-wide analysis supports environmental adaptations of two Pinus pinaster populations from contrasting habitats. BMC Genomics 16: 909 https://doi.org/10.1186/s12864-015-2177-x
CAMPINHOS, E. 1987. Propagación vegetativa de Eucalyptus spp. por enraizamento de estacas. In: Simposio sobre Silvicultura y Mejoramiento Genetico de Especies Forestales, Buenos Aires. Anais. Buenos Aires: CIEF. v.1, p.208- 214.
CUENCA B., GONZÁLEZ L., FERNÁNDEZ M. R. 2009. Micropropagación de genotipos adultos de Castanea sativa Mill. seleccionados por resistencia a Phytophthora cinnamomi. En: 5º congreso Forestal Español. Libro de Resúmenes. Ávila. España.
CIORDIA M., FEITO I., MAJADA J. 2015. Recursos fitogenéticos de castaño, cerezo y nogal. Tecnología Agroalimentaria 15: 28-36.
DANTAS A.K., MAJADA J., DANTAS F.K., DELATORRE C., GRANDA V., BALLEJO P., FEITO I. 2016. Rooting of minicuttings of Castanea sativa Mill. hybrid clones. Revista Árvore 40: 465-475.
DELATORRE C., FUENTE-MAQUEDA J.F., MEIJÓN M., FEITO I., BAIZÁN S., DANTAS A.K., MAJADA J., GRANDA V., RODRÍGUEZ A. 2013. Compuestos de naturaleza fenólica y actividad antioxidante de los brotes del pino (Pinus pinaster Aiton). Tecnología Agroalimentaria 13: 27-33
GARCÍA DEL BARRIO J. M., DE MIGUEL, J., ALÍA R., IGLESIAS S. 2001. Regiones de identificación y utilización de material forestal de reproducción. Serie cartográfica, Ministerio de Medio Ambiente.
GASPAR M.J, VELASCO T., FEITO I., ALÍA R., MAJADA J. 2013. Genetic variation of drought tolerance in Pinus pinaster at three hierarchical levels: A comparison of induced osmotic stress and field testing. PLoS ONE 8(11): e79094. https://doi.org/10.1371/journal.pone.0079094.
HARTMAN H., KESTER D. 1998. Propagación de plantas. Principios y prácticas. Trad. A. Marino. 3 ed. México, DF, Continental S.A.
LÓPEZ-GONZÁLEZ, G. 2001. Los árboles y arbustos de la Península Ibérica y Baleares. Ed Mundi-Prensa.
MACDONALD B. 1995. Principles of vegetative propagation, clonal selection and stock (Mother). Practical woody plant propagation for nursery grower. Portland Oregon. Timber Press.
MAJADA J., MARTÍNEZ_ALONSO C., FEITO I., KIDELMAN A., ARANDA I., ALIA R. 2011. Minicuttings: an effective technique for the propagation of Pinus pinaster Ait. New Forest 41(3): 399-412.
MARTÍNEZ-ALONSO C., KIDELMAN A., FEITO I., VELASCO T., ALÍA R., GASPAR M.J., MAJADA J. 2012. Optimization of seasonality and mother plant nutrition for vegetative propagation of Pinus pinaster Ait. New Forests 43: 651-663.
MEIJÓN M., FEITO I., ORAVEC M., DELATORRE C., WECKWERTH W., MAJADA J., VALLEDOR L. 2016. Exploring natural variation of Pinus pinaster Aiton using metabolomics: Is it possible to identify the region of origin of a pine from its metabolites? Molecular Ecology 25(4): 959-976.
MILLER G., MILLER D.D., JAYNES R.A. 1996. Chestnuts. In: Janick J. and Moore J.N. Fruit breeding. Ed. John Wiley and Sons, Inc, New York. pp: 99-124.
MONTERO G., CISNEROS O., CANELLAS I. 2003. Manual de selvicultura para plantaciones de especies productoras de madera de calidad. Madrid, Mundi-Prensa.
RITCHIE G.A. 1991. The commercial use of conifer rooted cuttings in forestry: A world overview. New Forests 5: 247-275.
RUSSELL K. 2003. EUFORGEN.Technical Guidelines for genetic conservation and use for wild cherry (Prunus avium). International Plant Genetic Resources Institute, Rome, Italy.
TITON M., XAVIER A., OTONI W. C. 2002. Dinâmica do enraizamento de microestacas e miniestacas de clones de Eucalyptus grandis. Revista Árvore 26 (6): 665-673.
WENDLING I., XAVIER A., GOMES J. M., PIRES I. E., ANDRADE H. B. 2000. Efeito do regulador de crescimento AIB na propagação de clones de Eucalyptus spp. Por miniestaquia. Revista Árvore 24 (2): 187-192.
WENDLING I., XAVIER A. 2001. Gradiente de maturação e rejuvenescimento aplicado em espécies florestais. Floresta e Ambiente 8(1):187-194.
XAVIER A., WENDLING I. 1998. Miniestaquia na clonagem de Eucalyptus. Viçosa, MG: SIF. (Informativo Técnico SIF, 11).
| Ficha Bibliográfica | |
|---|---|
| Título | Minipropagación de plantas forestales |
| Autor/es | ÂNGELO KIDELMAN DANTAS. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. kidelman3@hotmail.com JUAN MAJADA. Centro Tecnológico y Forestal de la Madera (CETEMAS). jmajada@cetemas.es MARTA CIORDIA. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. mciordia@serida.org ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales. Programa Forestal. ifeito@serida.org |
| Año Publicación | 2018 |
| Área | |
| Revista/Serie | Tecnología Agroalimentaria. Boletín informativo del SERIDA |
| Referencia | Nº 20. pp. 28 - 36 |
| Formato | |
| Depósito Legal | AS.-2.617/1995 |
| ISSN | 1135-6030 |
| ISBN | |
| Ver/Descargar | http://www.serida.org/publicacionesdetalle.php?id=7437&anyo= |