

Publicaciones SERIDA
El paisaje agrícola tradicional de Asturias limita la expansión de las plagas de roedores.
AITOR SOMOANO GARCÍA. Área de Cultivos Hortofrutícolas y Forestales. Programas de Investigación en Fruticultura. SERIDA. aitors@serida.org
JACINT VENTURA QUEJIA. Departament de Biología Animal, de BIología Vegetal i d' Ecología. Facultat de Biociènces. Universitat Autònoma de Barcelona. Área de REcerca en petit Mamìfers, Museu de Ciènces Naturals de Granollers. jacint.ventura.quejia@uab.cat
MARCOS MIÑARRO PRADO. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación en Fruticultura. SERIDA. mminarro@serida.org

La tendencia general a la intensificación de la agricultura desde mediados del siglo pasado ha provocado una disminución de la heterogeneidad del hábitat y un incremento de la presencia y la gravedad de los daños producidos por roedores perjudiciales para la agricultura, como el topillo campesino, Microtus arvalis, o la rata topera, Arvicola scherman (Figura 1) (Luque-Larena et al. 2013, Halliez et al. 2015). La rata topera está ampliamente extendida en Asturias, donde encuentra su hábitat idóneo en prados y plantaciones de frutales. Este roedor causa importantes pérdidas agrícolas al alimentarse de los cultivos y generar montones de tierra en superficie debido a su actividad excavadora. Se tiene constancia de que la rata topera es una especie perjudicial en esta región desde al menos el siglo XVII, cuando ya se señala su presencia en las llanuras de Villaviciosa y sus efectos dañinos en frutales y otros cultivos (Somoano 2020). Resultados recientes indican que, al menos en las zonas bajas de Asturias, la rata topera puede reproducirse durante todo el año mostrando un alto potencial reproductivo: una hembra puede llegar a producir hasta 28 crías en un solo año (Somoano et al. 2018).

La densidad de las poblaciones de rata topera experimenta oscilaciones muy notables, con años de elevada abundancia de individuos e importantes daños agrícolas asociados, seguidos de otros con densidades y perjuicios considerablemente menores. Cuando ocurren las explosiones demográficas (picos de población), estos roedores pueden dispersarse fácilmente en paisajes dominados por hábitats favorables, posibilitando que estos incrementos de densidad poblacional sucedan a gran escala. Es el caso de muchas regiones montañosas europeas dominadas por prados, donde esta especie puede extenderse en el territorio a medida que aumentan sus poblaciones (Halliez et al. 2015). El caso opuesto podría ser el paisaje agrícola asturiano, conformado por un mosaico de parcelas con diferentes usos del suelo separadas por setos o muros, y donde las manchas forestales son relativamente frecuentes. Se supone que una baja proporción de hábitats favorables compromete la dispersión y la colonización de nuevos hábitats. Para comprobar esta hipótesis hemos utilizado un enfoque que combina la genética de poblaciones y el estudio del paisaje a escala local en la Comarca de la Sidra, una zona tradicionalmente afectada por esta plaga. Según este planteamiento, si poblaciones relativamente próximas en el espacio son muy diferentes genéticamente es porque el paisaje está dificultando el contacto entre ellas. Y, al contrario, poblaciones en estrecho contacto serán genéticamente muy parecidas. El objetivo de este estudio fue determinar en qué medida el mosaico asturiano afecta a la capacidad de dispersión de la rata topera y minimiza la escala a la que suceden las explosiones demográficas.
Descripción del estudio
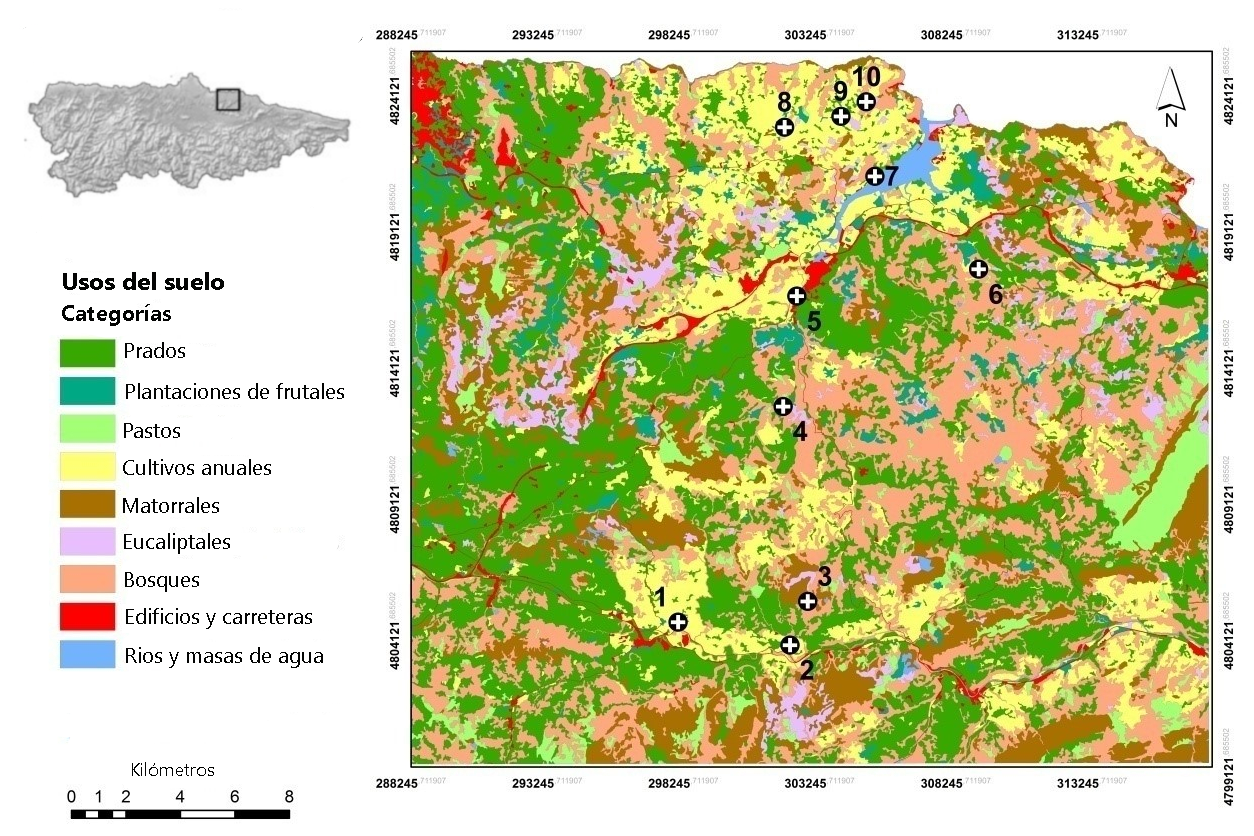
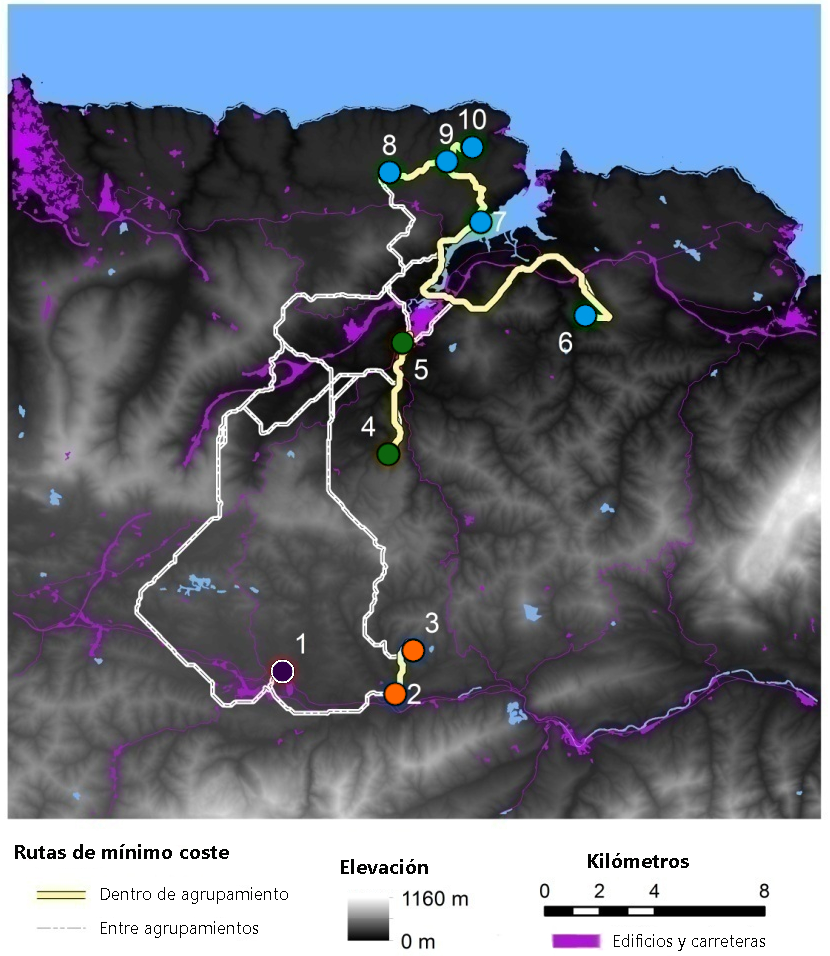
Se seleccionaron 10 plantaciones de manzano en los concejos de Villaviciosa y Nava, con una distancia entre sitios que osciló entre 1,1 y 20,5 km. En cada plantación se capturaron entre 11 y 18 ejemplares de rata topera (137 en total) para evaluar la variación genética a nivel de población y de individuo. Los análisis genéticos se realizaron a partir del ADN genómico. Cada muestra se analizó mediante amplificación por PCR de 12 loci microsatélites divididos en dos paneles multiplex (para más detalle metodológico ver Somoano et al. 2022). Tras evaluar los genotipos resultantes, se determinaron las diferencias genéticas entre individuos y la estructura genética poblacional en este territorio agrícola. Es decir, se estudió cuál era el grado de parentesco entre los individuos y su representación en el terreno. A su vez, se caracterizaron la cobertura y el uso del suelo en el área de estudio mediante sistemas de información geográfica y se desarrolló un modelo de resistencia a la dispersión según la habitabilidad y la permeabilidad de los hábitats para la rata topera (hábitats favorables: prados, plantaciones de frutales, pastos; hábitats desfavorables: matorrales, eucaliptales, bosques, carreteras) (Figura 2). De esta manera, además de las distancias en línea recta (llamadas distancias euclidianas) se obtuvieron unas distancias que tienen en cuenta la dificultad que ofrece cada hábitat a ser atravesado por la especie y que se conocen como distancias de resistencia. Por último, también se tuvo en cuenta la idoneidad del paisaje que separaba las poblaciones, que se calculó como la proporción de hábitats favorables para la rata topera respecto al total.
Estructura genética: ¿cómo están relacionadas las poblaciones?
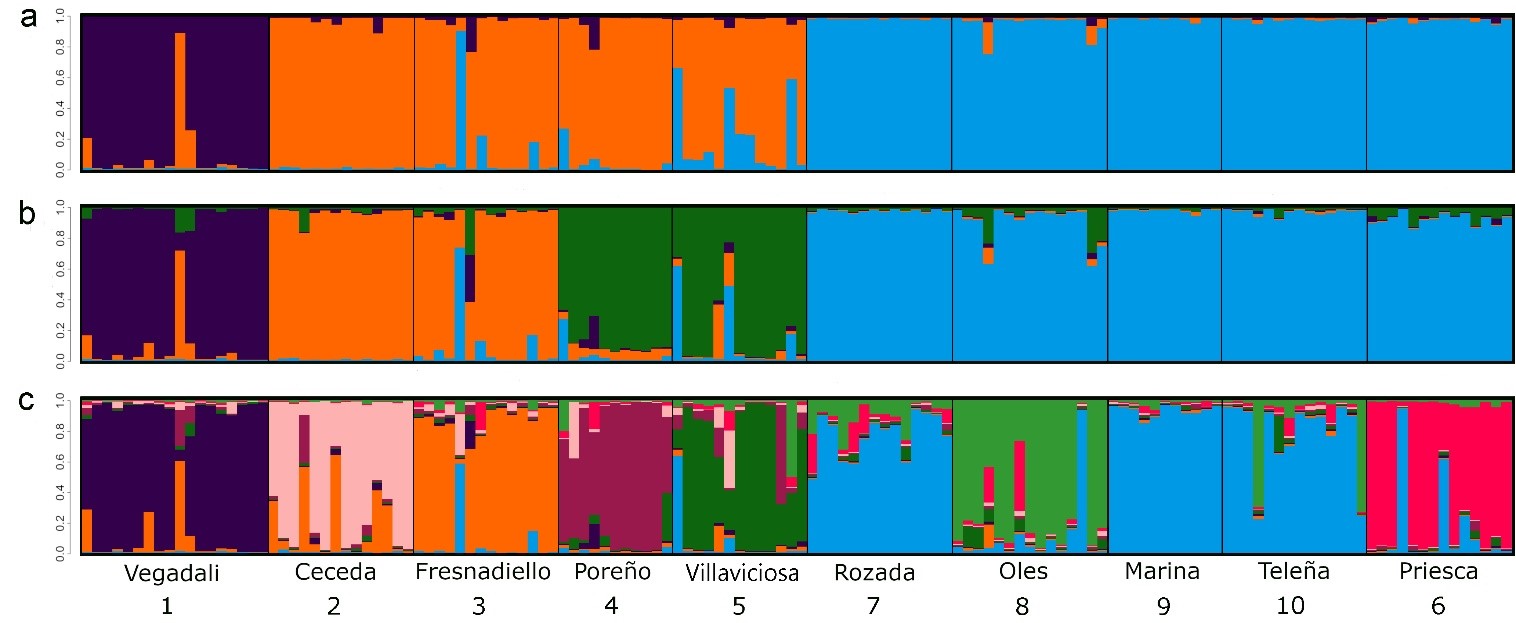
La estimación total de la distancia genética mostró un valor medio de 0,168 (rango: 0,128 - 0,219), lo que refleja una alta diferenciación genética o, lo que es lo mismo, un bajo grado de parentesco entre las poblaciones analizadas. El número de agrupaciones genéticas entre individuos (K), teniendo en cuenta las distancias (software Geneland), dio como resultado más probable K = 4 en todas las repeticiones, con una distribución de las poblaciones en dirección norte-sur (Figura 3). Según esto, se distinguirían 4 grupos de poblaciones distintas (representadas por diferentes colores en la Figura 3). Sin tener en cuenta la distancia espacial (software Structure), el análisis indicó la máxima probabilidad para K = 3 (Figura 4a) y K = 8 (Figura 4c). Ambas aproximaciones coinciden en una distribución significativa de los individuos en 4 grupos (K = 4 ,Figura 3b). No obstante, K = 8 reflejó también una agrupación altamente consistente, donde se observa una subdivisión de los grupos poblacionales mayores en grupos más pequeños. Estos resultados indican que en la zona de estudio podrían distinguirse, según la metodología empleada, desde 3 hasta 8 agrupaciones de rata topera con contacto limitado entre sí.
El paisaje influye en la estructura genética
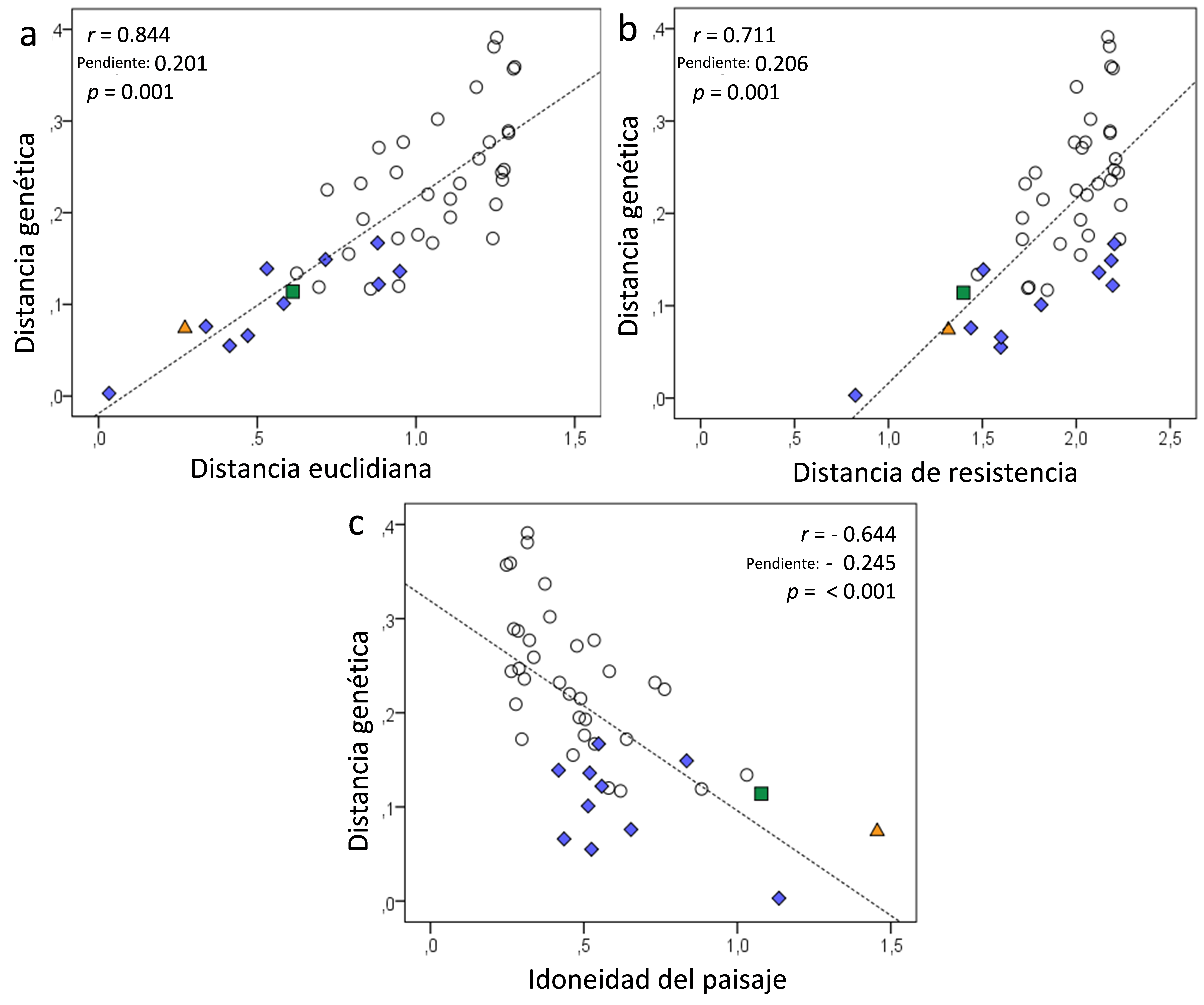
La relación entre la distancia euclidiana entre poblaciones y su diferenciación genética fue positiva y altamente significativa, lo que demuestra un fuerte patrón de aislamiento por distancia (Figura 5a), que corresponde a una elevada acumulación de variación genética asociada a una dispersión limitada. Cuando se consideraron las distancias de resistencia a la dispersión, también hubo una relación positiva altamente significativa y, por tanto, un patrón de aislamiento debido a esa resistencia (Figura 5b). Todos estos resultados indican que cuanto más separadas en el espacio estén las poblaciones de ratas toperas y más dificultades tengan éstas para atravesar los distintos hábitats, mayor será el aislamiento y mayores las diferencias genéticas entre poblaciones. Asimismo, se observó un porcentaje relativamente bajo de hábitats favorables para la especie en el área de estudio (31,6 % ± 10,0 desviación estándar), incluso dentro de los agrupamientos (idoneidad del paisaje: 24,4; 49,0 y 53,4% respectivamente en las agrupaciones de norte a sur para K = 3). De hecho, los datos de idoneidad del paisaje mostraron una relación inversa significativa con la diferenciación genética entre poblaciones (Figura 5c), es decir, el aislamiento entre estas fue mayor cuanto menos favorable era el paisaje que las separaba. Esto demuestra que un paisaje en mosaico, con abundancia de hábitats desfavorables para la rata topera, supone un obstáculo para su dispersión y es coherente con la observación de que los individuos dentro de las agrupaciones estaban más emparentados entre sí que cuando se consideraban individuos de cualquier población escogidos al azar. Esta relación de parentesco se mantuvo sólo hasta 2800 m de distancia, a diferencia de lo que ocurre en paisajes homogéneos, donde el parentesco se mantiene hasta los 30 km (Somoano et al. 2022).
Conclusiones
Con un enfoque de genética de poblaciones se ha probado la hipótesis de que paisajes heterogéneos comprometen la colonización de nuevos hábitats por parte de las plagas de roedores, lo que se refleja aquí en la estructura genética de la rata topera a escala local. La alta diferenciación genética entre poblaciones indicó un patrón jerárquico de hasta ocho agrupamientos potencialmente relevantes, independientemente de la distancia a la que se encontrasen dichas poblaciones. De acuerdo con nuestra hipótesis, la configuración del paisaje agrícola tradicional de Asturias puede considerarse un factor clave para limitar el avance de las poblaciones de la rata topera. La baja conectividad entre hábitats adecuados y la relativa baja proporción de estos hacen que este paisaje sea poco permeable a este roedor, y que la colonización de nuevos territorios se vea limitada por dificultades de dispersión y el aumento de la mortalidad asociada a la depredación (Somoano et al. 2022).
Implicaciones para el control poblacional de la plaga
La variación genética observada parece indicar que la dinámica poblacional de la rata topera consiste en un conjunto de subpoblaciones (aquí agrupamientos) separadas en el espacio que mantienen solo un cierto grado de interacción; se trataría de una metapoblación. Los agrupamientos determinados en el presente estudio pueden considerarse, por tanto, unidades demográficas en gran medida independientes, cuyo tamaño depende principalmente de las tasas locales de natalidad y mortalidad, más que del intercambio de individuos entre poblaciones próximas. En paisajes agrícolas homogéneos las explosiones demográficas de rata topera se producen a escala regional debido a la dispersión sin obstáculos, y por tanto el manejo de esta plaga debe abordarse a gran escala. En cambio, la estructura genética de esta especie en el paisaje agrícola heterogéneo de Asturias indica que sus explosiones demográficas pueden tener lugar a escala de subpoblación y el manejo, por tanto, puede abordarse a una escala menor y de manera independiente del de zonas relativamente próximas. Así, una proporción global de hábitats favorables por debajo del 35% podría ser una limitación para la dispersión de las ratas toperas más allá de los 2,8 km. Nuestros resultados apoyan empíricamente el valor intrínseco de los paisajes complejos para modular las densidades poblacionales de especies que dependen de las características del hábitat, como son los roedores perjudiciales para la agricultura.
Agradecimientos
A los propietarios que nos permitieron capturar roedores en sus pomaradas. Esta investigación ha sido financiada por los proyectos FICYT PC10-52 (Gobierno del Principado de Asturias, FEDER, Caja Rural de Gijón, CADAE y AACOMASI), RTA2013-00039-C03-01 y RTA2017-00051-C02-01 (INIA, Ministerio de Economía y Competitividad y FEDER), PCTI 2021-2023 (GRUPIN: IDI-2021-000102, Gobierno del Principado de Asturias y FEDER), y el programa PTA2020-AEI (AS).
Referencias bibliográficas
HALLIEZ, G., RENAULT, F., VANNARD, E., FARNY, G., LAVOREL, S., GIRADOUX, P. 2015. Historical agricultural changes and the expansion of a water vole population in an Alpine valley. Agriculture, Ecosystems and Environment. 212: 198–206.
LUQUE-LAERNA, J.J., MOUGEOT, F. VINEULA, J. ,JAREÑO, D., ARROYO, L., LAMBIN, X., ARROYO, B. 2013. . Recent large-scale range expansion and outbreaks of the common vole (Microtus arvalis) in NW Spain. Basic and Applied Ecology. 14: 432-441.
SOMOANO, A., VENTURA, J. , MIÑARRO, M. 2018. Reproducción continua de la rata topera en pomaradas asturianas. Tecnología Agroalimentaria. 20: 22 - 27.
SOMOANO, A. 2020. El papel de la rata topera (Arvícola scherman) como plaga agrícola en el noroeste de España: ¿desde cuándo? Galemys 32: 61–63.
SOMOANO, A. , BASTOS-SILVEIRA, C., VENTURA, J. MIÑARRO, M., HECKEL, G. 2022 Bocage landscape restricts the gene flow of pest vole populations. Life 12: 800.
| Ficha Bibliográfica | |
|---|---|
| Título | El paisaje agrícola tradicional de Asturias limita la expansión de las plagas de roedores. |
| Autor/es | AITOR SOMOANO GARCÍA. Área de Cultivos Hortofrutícolas y Forestales. Programas de Investigación en Fruticultura. SERIDA. aitors@serida.org JACINT VENTURA QUEJIA. Departament de Biología Animal, de BIología Vegetal i d' Ecología. Facultat de Biociènces. Universitat Autònoma de Barcelona. Área de REcerca en petit Mamìfers, Museu de Ciènces Naturals de Granollers. jacint.ventura.quejia@uab.cat MARCOS MIÑARRO PRADO. Área de Cultivos Hortofrutícolas y Forestales. Programa de Investigación en Fruticultura. SERIDA. mminarro@serida.org |
| Año Publicación | 2022 |
| Área | Sanidad Animal. |
| Revista/Serie | Tecnología Agroalimentaria. Boletín del SERIDA. |
| Referencia | Nº27. Pags 24-29. |
| Formato | |
| Depósito Legal | 2.617/1995 |
| ISSN | 1135-6030 |
| ISBN | |
| Ver/Descargar | |