

Resultado Proyecto
Patología vegetal. Enfermedad en castaño
Duración: 2019-2019.
Equipo investigador
Ana J. González Fernández SERIDA
Entidades Colaboradoras
Laboratorio Oficial de Sanidad Vegetal del Principado de Asturias,
Marta Ciordia Ara. SERIDA (Programa Investigación Forestal)
Avance de resultados
Enfermedad en castaño:
En mayo se detectaron síntomas de necrosis apical que avanza hacia abajo con daño vascular en un ensayo de castaño en el que las plantas podían llegan a morir.
Las muestras se analizaron para identificar la presencia de potenciales hongos y bacterias patógenos. Mediante métodos moleculares se identificó la bacteria Staphylococcus pasteuri; sin embargo, y como prevención, se enviaron extractos de la muestra al Laboratorio de Sanidad Vegetal del Principado de Asturias que descartó la presencia de Xylella fastidiosa.
En cuanto a los hongos, se pudieron identificar, por una parte Alternaria tenuis, que no es considerado patógeno y, por otra, Neopestalotiopsis clavispora, Diaporthe eres y Diaporthe foeniculina. Estas tres últimas especies se han descrito como patógenas de varias especies aunque no en castaño, por lo que se van a realizar ensayos de patogenicidad en planta para aclarar su papel en la enfermedad observada, en colaboración con el Programa de Investigación Forestal.
Bioensayo de Neopestalotiopsis zimbabwana frente a Cryphonectria parasitica
Se amplió el estudio previo realizando el ensayo de la cepa AS-860 de N. zimbabwana frente a las cepas de C. parasitica AS-26 y AS-766 en sus variantes virulenta e hipovirulenta. Se ha visto en estos ensayos que los resultados de ambos tipos de cepas son muy similares y se apunta a este hongo como un posible agente de biocontrol.
Bioensayo de producción de lipodepsipéptidos en Pseudomonas syringae
Se ha utilizado la cepa de Rhodotorula mucilaginosa CECT 11016 frente a 30 cepas de P. syringae, en el medio PGNaClA (peptona-glucosa-cloruro sódico-agar). De las 30 cepas ensayadas, 18 se habían asignado al patovar syringae de acuerdo con sus características fenotípicas y la amplificación de las bandas correspondientes a syrB, syrD y sypA. Sin embargo, sólo 14 de los 18 han inhibido el crecimiento de R. mucilaginosa, lo que se podría explicar por deficiencias en la biosíntesis de las toxinas o fallos en la expresión. Once cepas no asignadas al pv syringae (con sypA o sypA y syrD amplificados) inhibieron el crecimiento de la levadura lo que podría corresponder a la producción de siringopeptina o de otras toxinas como siringotoxina y siringostatina que también se han descrito en P. syringae. Una única cepa no inhibió el crecimiento de la levadura y no había amplificado ninguno de los genes testados.
Con estos resultados no podemos considerar que este bioensayo sea un método a incluir en los protocolos de identificación de P. syringae pv syringae.

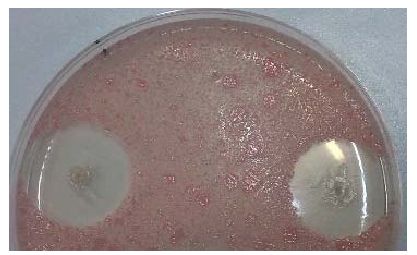
Figura 1.- A la izquierda, sintomatología del castaño analizado. A la derecha, inhibición del crecimiento de R. mucilaginosa por P. syringae, a ambos lados de la placa se observan los halos de inhibición y en el centro arriba, ausencia de la misma.